Рис. 1. Ноги ракообразных и насекомых. Вверху: бокоплав Parhyale и его третья грудная нога. Внизу: клоп Oncopeltus и его вторая грудная нога. Изображение из обсуждаемой статьи в Nature Ecology and Evolution
Эксперименты с генетически модифицированными ракообразными (бокоплавами Parhyale) позволили американским биологам подтвердить и развить старую гипотезу о том, что крылья насекомых произошли от боковых отростков (экзитов) ног ракообразных. Между члениками ног раков и насекомых удалось установить четкое соответствие по принципу «один к одному»: шесть члеников ног насекомых соответствуют шести дистальным (самым дальним от тела) членикам ног ракообразных. Два базальных членика ноги ракообразных (седьмой и восьмой, кокса и прекокса) у насекомых вошли в состав стенки тела, образовав ее боковую часть — плейрит. Крылья насекомых, по-видимому, представляют собой экзиты восьмого членика (прекоксы).
Происхождение крыльев насекомых — давняя научная загадка. Ископаемая летопись не сохранила внятных переходных форм между первичнобескрылыми и крылатыми насекомыми, поэтому ученым приходится полагаться лишь на данные таких дисциплин, как сравнительная анатомия и эволюционная биология развития.
В основном обсуждаются две версии. Согласно первой, крылья произошли от экзитов — отростков проксимальной (базальной, ближайшей к телу) части примитивной конечности древних ракообразных — предков насекомых. Это подтверждается, в частности, тем, что некоторые ключевые гены — регуляторы развития сходным образом экспрессируются в экзитах ракообразных и крыльях насекомых в ходе онтогенеза (N. Shubin et al., 1997. Fossils, genes and the evolution of animal limbs).
Но как может быть крыло частью ноги, если между настоящей ногой и крылом у насекомых находится боковая часть стенки тела — плейрит, вроде бы совсем не похожий на конечность? Дело в том, что многие специалисты по сравнительной анатомии и эмбриологии членистоногих уже давно, начиная аж с конца XIX века, высказывали идею, что плейриты грудного отдела насекомых образовались из базальной части ноги, возможно, из базального членика — субкоксы. Это называют «субкоксальной теорией». Предполагается, что этот членик, отсутствующий у современных насекомых, но имевшийся у предков, встроился в стенку тела и стал плейритом. Или, может быть, это произошло даже не с одним, а с двумя базальными члениками — субкоксой-1 и субкоксой-2 (Y. Kobayashi, 2017. Formation of Subcoxae-1 and 2 in Insect Embryos: The Subcoxal Theory Revisited). В таком случае логично, что экзит — уплощенный вырост базального членика, встроившегося в стенку тела — оказался как раз там, где у насекомых находятся крылья: в самой верхней части плейрита, на границе со спинным щитком (тергитом).
Здесь можно еще вспомнить (хоть это и не имеет прямого отношения к статье, о которой пойдет речь), что примитивная конечность древних членистоногих могла развиться путем слияния двух исходно независимых конечностей, вентральной и дорзальной (см.: Новооткрытый родственник аномалокариса проливает свет на происхождение конечностей членистоногих, «Элементы», 16.03.2015). И тогда получается, что предполагаемое разделение единой конечности на крыло и ходильную ногу у крылатых насекомых в каком-то смысле было возвратом к очень древнему предковому состоянию, а шипы галлюцигении (см.: Палеонтологи выяснили, как была устроена голова галлюцигении, «Элементы», 26.06.2015) могут в итоге оказаться гомологичными крыльям насекомых.
Вторая версия предполагает, что крылья развились не из отростков ног — экзитов, а из выростов стенки тела, не связанных с ногами, — так называемых паранотальных выростов. Такие выросты по краям тергитов встречаются у первичнобескрылых насекомых, причем «крыловые» гены в них тоже исправно экспрессируются. У предков крылатых насекомых (Pterygota) они могли служить, например, для планирования при прыжках с ветки на ветку. В этом случае — если крылья никогда не были частью ноги — сходство генетической регуляции развития крыльев и экзитов следует трактовать как результат «коопции» регуляторных генных сетей. То есть гены, у предков отвечавшие за развитие придатка ноги, позднее были привлечены для выполнения новой функции — регуляции развития паранотальных выростов, позднее превратившихся в крылья. В этом случае уже нельзя говорить о строгой гомологии крыльев и экзитов, хотя и «подлинным эволюционным новшеством», развившимся с нуля, назвать крылья тоже не получится. Здесь полагается рассуждать о так называемой глубокой гомологии (N. Shubin et al., 2009. Deep homology and the origins of evolutionary novelty), о размытости понятия «эволюционное новшество», о непригодности формулировки «новшество начинается там, где кончается гомология» — в общем, об отсутствии грани между подлинным новшеством и классическим дарвиновским «наследованием с изменениями» (descent with modification; хороший недавний обзор на эту тему: D. M. Linz et al., 2020. From descent with modification to the origins of novelty).
В последнее время популярны также комбинированные версии, согласно которым крыло насекомого — это отчасти видоизмененный экзит древней конечности ракообразного предка, а отчасти — паранотальный вырост, то есть «новая» структура, не связанная с ногами древних ракообразных (N. Niwa et al., 2010. Evolutionary origin of the insect wing via integration of two developmental modules).
Пытаясь разобраться в этом запутанном вопросе, американские биологи провели серию генетических экспериментов с разноногим рачком (бокоплавом) Parhyale hawaiensis. Главной задачей исследования было понять, как соотносятся друг с другом членики конечностей ракообразных и насекомых. Этот вопрос имеет огромное значение для понимания происхождения крыльев. Ведь если бы удалось показать, что самый базальный членик ноги насекомого (кокса) гомологичен не самому базальному членику ноги ракообразного, это было бы сильным аргументом в пользу субкоксальной теории (согласно которой базальная часть предковой ноги встроилась в стенку тела, став плейритом). Забегая вперед, заметим, что именно это авторам и удалось показать.
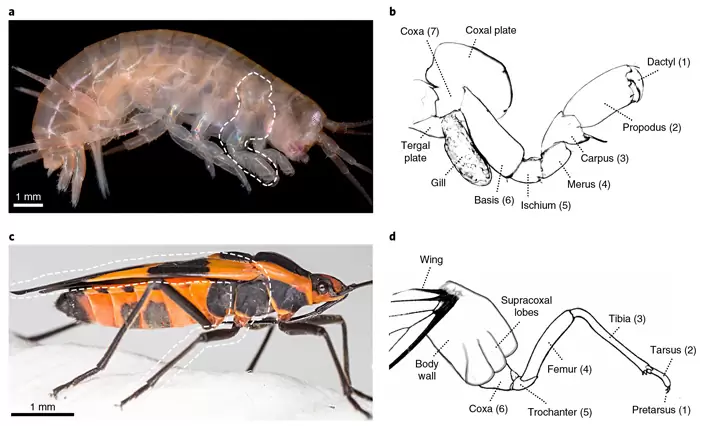
Грудные ноги бокоплава Parhyale состоят из семи члеников (рис. 1). На самом базальном (седьмом) членике, который тоже называется коксой, находятся два отростка — экзита: жаберный отросток и уплощенная коксальная пластинка (coxal plate). Для насекомых типичны ноги из шести члеников, как у показанного на рис. 1 клопа Oncopeltus.
Чтобы разобраться в гомологии члеников ног, ученые при помощи CRISPR/Cas9 поочередно выводили из строя те гены рачка, которые у дрозофилы отвечают за разметку конечности вдоль проксимально-дистальной оси и формирование члеников. Исследовали пять таких генов: Distalless (Dll), Sp6–9, dachshund (dac), extradenticle (exd) и homothorax (hth). Ранее было показано, что у насекомых (в основном такие исследования проводились на дрозофиле, реже — на других модельных видах) при отключении каждого из этих генов пропадают строго определенные членики ног. Поэтому для того, чтобы установить соответствие между члениками ног насекомых и ракообразных, нужно было посмотреть, какие членики пропадут при отключении этих генов у бокоплава.
Результаты получились красивые и показательные (рис. 2).
Фенотипические эффекты отключения каждого из пяти генов у бокоплава в точности совпали с таковыми у насекомых. При этом между члениками устанавливается четкое соответствие «один к одному», начиная с самого дистального (первого) и вплоть до шестого, который у насекомых называется коксой, а у раков — базисом.
Например, у насекомых при отключении Dll не развиваются членики 1–5, и у Parhyale результат оказался точно таким же (рис. 2, b). При отключении Sp6-9 у насекомых, как и у бокоплава, исчезают членики 1–6 (рис. 2, d), и так далее. Различия минимальны: например, у дрозофилы ген dac необходим для развития члеников 3–5 и отчасти членика 2, а у Parhyale — только для члеников 3–5.
Эти эксперименты показали, что членики 1–6, из которых состоит нога насекомого, по-видимому, гомологичны членикам 1–6 ноги бокоплава. Кокса насекомых соответствует не коксе, а базису ракообразных. Но вот членика 7 (кокса ракообразных) у насекомых нет. Точнее, его нет в составе ноги. Но ведь мы помним, что, согласно «субкоксальной теории», он мог стать частью стенки тела (плейрита). С другой стороны, насекомые могли и просто утратить этот членик.
Чтобы выяснить судьбу седьмого членика, исследователям пришлось искать гены, по рисунку экспрессии которых можно надежно отличить основание ноги от стенки тела. Самыми удобными маркёрами оказались гены pannier (pnr), маркирующий дорзальную стенку тела (которая уж точно не является бывшей ногой), и araucan (ara). Рисунок экспрессии ara у эмбрионов бокоплава и жука Tribolium оказался удивительно похожим и очень показательным. Жук в этих экспериментах представлял насекомых, потому что у личинок дрозофил нет ног, а у личинок жуков — есть. В основании каждой конечности и у эмбриона жука, и у эмбриона бокоплава обнаружились три области экспрессии ara. Эти области на рис. 3 выделены зеленым цветом и отмечены тремя разными стрелками.
Первая, самая дистальная область экспрессии ara (на рисунке она справа, отмечена стрелками с «древком») приурочена к шестому членику ноги. У рачка это базис, у жука — кокса.
Вторая область (отмечена незакрашенными стрелками без древка) маркирует у рачка основание (проксимальную часть) седьмого членика — коксы ракообразных. Эта область экспрессии находится на границе седьмого членика с тем, что у взрослого рачка станет стенкой тела, а у эмбриона до боли напоминает восьмой членик ноги. Кстати, у многих ракообразных нога действительно имеет восьмой членик, самый базальный — прекоксу. У Parhyale он, судя по всему, инкорпорирован в стенку тела.
Третья область экспрессии ara (закрашенные стрелки без древка) у рачка приурочена к основанию восьмого членика. Из эмбриональных тканей, расположенных между второй и третьей областями экспрессии ara, то есть из восьмого членика эмбриональной ноги, у взрослого рачка развивается часть стенки тела с характерным плоским отростком — тергальной пластинкой (Tp на рис. 3). Такие отростки есть у многих ракообразных, и в них экспрессируются «крыловые» гены. Авторы делают обоснованный вывод, что тергальная пластинка — это экзит восьмого членика ноги. На седьмом членике есть похожий отросток-экзит — коксальная пластинка (Cp на рис. 3), а также жабра (G), которую можно интерпретировать как еще один экзит седьмого членика.
Что касается жука, то у него вторая область экспрессии ara тоже находится на границе эмбриональных структур, выглядящих как седьмой и восьмой членики ноги, а третья — у основания восьмого членика. Но у жука, как мы знаем, в ногах нет ни седьмого членика, ни восьмого. Из эмбриональных зачатков члеников 7 и 8 развивается плейрит — боковая стенка тела. У некоторых насекомых на плейрите есть уплощенный выступ (или несколько выступов, как у клопа на рис. 1; они подписаны «Supracoxal lobes»). Эти выступы напоминают коксальные пластинки ракообразных. Авторы полагают, что они действительно гомологичны коксальным пластинкам и являются экзитами седьмого членика, встроенного в стенку тела (рис. 4). Ну а из зачатка членика 8, где у эмбриона жука находится дыхальце (Spiracle на рис. 3), у взрослого жука на втором и третьем грудных сегментах вырастут самые замечательные экзиты — крылья (задние летательные и передние — жесткие элитры).
Авторы приводят в статье ряд дополнительных анатомических, эмбриологических и генетических аргументов в пользу своей трактовки, отчасти основанных на собственных экспериментах, отчасти — на литературных данных. В частности, они упоминают, что у насекомых на плейрите иногда заметен шов — рудиментарное сочленение между седьмым и восьмым члениками ноги, влившимися в стенку тела. В этом сочленении у жука Tribolium зарегистрирована экспрессия гена Serrate, которая маркирует суставы ног у насекомых.
Главные выводы исследования суммированы на рис. 5. В целом надо признать, что концепция получилась хоть и не бесспорная (специалистам предстоит еще много работы, прежде чем вопрос о происхождении крыльев насекомых можно будет считать закрытым), но, безусловно, красивая.
Ну а противопоставление двух теорий происхождения крыльев, о котором говорилось в начале новости, скорее всего, представляет собой мнимую дихотомию. Ведь экзиты восьмого членика предковой ноги не сразу стали крыльями. У непосредственных предков Pterygota, первичнобескрылых насекомых, они, очевидно, были обычными паранотальными выростами, похожими на тергальные пластинки Parhyale. Например, у первичнобескрылого насекомого Pedetontus unimaculatus (семейство Machilidae) паранотальные выросты на грудных сегментах действительно похожи на тергальные пластинки Parhyale, и в их развитии тоже задействованы «крыловые» гены.
Исследование: Heather S. Bruce & Nipam H. Patel. Knockout of crustacean leg patterning genes suggests that insect wings and body walls evolved from ancient leg segments // Nature Ecology & Evolution. 2020. DOI: 10.1038/s41559-020-01349-0.
Текст: Александр Марков | Источник: elementy
48 человек, регулярно курящих каннабис, участвовали в двойном слепом плацебо-контролируемом эксперименте с четырьмя разными дозами,… Читать далее
Женщины во время ходьбы в темноте активнее сканируют обстановку вокруг. По мнению исследователей, такое внимание… Читать далее
Это случай из клинической практики про эффект ноцебо. 26-летний мужчина обратился в регистратуру отделения неотложной… Читать далее
Исследователи могут вызвать конкретный тип смеха точечным разрядом тока. Когда мы искренне хохочем над отличной… Читать далее
Артроскопия коленного сустава при дегенеративных заболеваниях коленного сустава считается одной из самых распространённых ортопедических операций.… Читать далее
Популярная идея о том, что микротравмы ответственны за рост мышц, появилась не на пустом месте.… Читать далее




